Cannabinoids have been used in Eastern medicine for many years as pain relievers. Δ9-tetrahydrocannabinol (THC), the major psychoactive compound in marijuana and hashish, has been shown to interact with two specific cannabinoid receptors: cannabinoid receptor 1 (CB1 Receptor or CB1R) and cannabinoid receptor 2 (CB2 Receptor or CB2R). The cannabinoid receptors can be distinguished by their amino acid sequences, signaling mechanisms, and tissue distributions. Both receptors belong to the G-protein-coupled receptor (GPCR) superfamily. Alomone Labs is pleased to offer antibodies to both receptors. This paper cites the use of these antibodies in various scientific papers.
Introduction
Endogenous cannabinoids are a family of molecules derived from arachidonic acid that signal through CB1 and CB2 receptors. The receptors belong to the superfamily of seven transmembrane-spanning regions; G-protein coupled receptors (GPCR)11. The cannabinoid receptor subtype-1 (CB1) is the most highly expressed GPCR in the central nervous system and is present at presynaptic terminals and axonal fibers in many brain areas, where it is believed to inhibit neurotransmitter release following agonist binding4.
Cannabinoids mediate anti-nociception via actions in a number of regions throughout the central nervous system. Systemic and intracerebroventricular administration of cannabinoid agonists both produce analgesia8. Microinjection of cannabinoids into several brain regions, including the rostral ventromedial medulla (RVM) and periaqueductal grey (PAG), also produces anti-nociception10.
CB1 Receptors
Endocannabinoid signaling has been shown to mediate synaptic plasticity by retrogradely inhibiting presynaptic transmitter release in several systems13. Gonzalez-Islas et al. found that endocannabinoids act tonically to regulate AMPA miniature postsynaptic current (mPSC) frequency in embryonic motor circuits of the chick spinal cord5. Indeed, CB1 expression was detected by western blotting embryonic chick spinal cord homogenates using Anti-Cannabinoid Receptor 1 (extracellular) Antibody (#ACR-001). The specificity of the antibody was shown by preincubating lysates with the negative control antigen prior to the antibody incubation (Figure 1). Furthermore, strong postsynaptic depolarizations also induced a short-lived endocannabinoidmediated suppression of mEPSC frequency. They determined that chronically inhibiting CB1 in vivo increases the frequency of spontaneous network activity (SNA) leading to a compensatory reduction in excitatory GABA and AMPA miniature postsynaptic current (mPSC) amplitude. The results suggest a role for tonic signaling of endocannabinoids as a potential mechanism to regulate the level of SNA, which is known to be critical for synaptic maturation in the embryonic spinal cord.
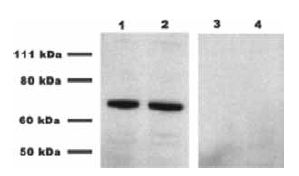
Western blot analysis of E10 chick embryo spinal cord using Anti-Cannabinoid Receptor 1 (extracellular) Antibody (#ACR-001). Lanes 1 and 2 depict the spinal cord lysate incubated with the antibody, while lanes 3 and 4 show lysates preincubated with the negative control antigen.
Adapted from reference 5 with permission of the Society for Neuroscience.
CB1 receptors are present in the areas of the brain involved in modulation of nociception3. Jennings, E.A. et al. examined the cellular actions of cannabinoids on neurons in the substantia gelatinosa (SG) of the spinal trigeminal nucleus pars caudalis, using whole-cell and perforated patch recording in brain slices. The recordings were performed in the presence of Tetrodotoxin citrate (#T-550)9. This study demonstrates that cannabinoids acting via presynaptic CB1 receptors inhibit both GABAergic and glycinergic synaptic transmission from presumptive interneurons, without altering non-NMDA receptor mediated glutamatergic transmission from primary afferents in the medullary dorsal horn. In addition, cannabinoids had no postsynaptic effects on medullary SG neurons. The present results suggest that cannabinoids are hyperalgesic at the level of the medullary dorsal horn because of their selective inhibition of GABAergic and glycinergic transmission.
Several studies have reported changes in intracellular Ca2+ concentration ([Ca2+]in) by cannabinoid compounds in both neuronal and non-neuronal preparations15. The effects of cannabinoid receptor stimulation on membrane currents in single cells from the Syrian hamster vas deferens cell line DDT1MF-2 were investigated using whole cell patch clamp1. The goals of the study were to see whether the increase in [Ca2+]incaused by CB1 receptor stimulation evoked a Ca2+-dependent K+ current, to establish the source of the increase in [Ca2+]in, and to determine the signaling pathway(s) leading to Ca2+ mobilization via CB1. The synthetic cannabinoid receptor agonist CP55,940 evoked a concentration-dependent outward current that was inhibited by a CB1-selective ligand but not by a CB2-selective ligand, suggesting that the response was mediated by CB1 receptors. Inhibition of mitogen-activating protein kinase (MAP kinase) and adenylyl cyclase blocked the outward Ca2+ current evoked. In addition, Iberiotoxin (#STI-400) or Charybdotoxin (#STC-325) inhibited or completely abolished the cannabinoid induced outward current. These data suggest that CB1 receptor stimulation in DDT1MF-2 cells leads to activation of a large conductance Ca2+-dependent K+ channel through a G-protein-mediated increase in [Ca2+]in, for which both inhibition of adenylyl cyclase and activation of MAP kinase are required. In addition, the cannabinoid-induced increase in [Ca2+]in is likely to arise from capacitive Ca2+ entry.
In central neurons, the cell-surface distribution of CB1 is highly polarized toward axons and is associated with synaptic terminals, where it is well positioned to modulate neurotransmitter release. Recent work investigated the mechanisms responsible for CB1 receptor axonal polarization in hippocampal neurons12. In experiments, N-terminally tagged enhanced green fluorescent protein (GFP)-CB1 chimeras demonstrate that CB1 receptors are delivered to both axonal and somatodendritic plasma membranes but that surface polarity is achieved by selective, constitutive endocytosis/basal receptor removal. Inhibiting endocytosis by transfecting cells with dominant-negative Dynamin-1 constructs (a protein involved in endocytosis), caused endogenous CB1 receptors to remain at the cell surface and not reposition in a polarized manner. This effect was observed in live rat hippocampal neurons using Anti-Cannabinoid Receptor 1 (extracellular) antibody which monitored cell surface CB1 receptors. Inhibiting CB1 receptor had no effect on the axonal polarization of the receptor. However, treating cells with a CB1 agonist led to a significant reduction of cell surface labeling of the receptor and an increase in CB1 labeling in somatodendritic regions (Figure 2)12. These data suggest that domain-specific endocytosis regulates the functional polarization of CB1 receptors.
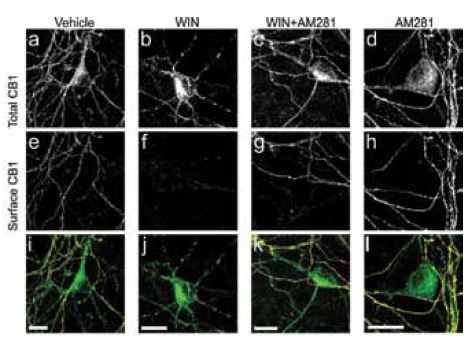
Immunocytochemical staining of rat hippocampal neurons left untreated (vehicle), treated with CB1 agonist (WIN), CB1 antagonist (AM281) and both (WIN+AM281). Extracellular labeling of cells using Anti-Cannabinoid Receptor 1 (extracellular) Antibody (#ACR-001) shows significant decrease in cell surface CB1 receptors (panel f) following Win55212-2 (WIN) treatment as opposed to AM281 treatment.
Adapted from reference 12 with permission of The American Society of Pharmacology and Experimental Therapeutics.
Several studies have shown that chronic liver disease is associated with the upregulation of endocannabinoids and their receptor, CB114. Chen et al. suppressed CB1 expression by siRNA and investigated its effect on hepatic fibrosis in vitro and in vivo2. The results demonstrate that siRNA significantly inhibited CB1 expression, as shown in western blot analysis using Anti-Cannabinoid Receptor 1 (extracellular) antibody and suppressed proliferation and extracellular matrix production in hepatic stellate cells (HSCs). Furthermore, CB1 knockdown significantly improved dimethylnitrosamine induced hepatic fibrosis, associated with the decreased expression of mesenchymal cell markers smooth muscle α-actin, vimentin and snail, and the increased expression of epithelial cell marker E-cadherin. The results suggest that CB1 is implicated in hepatic fibrosis and selective suppression of CB1 by small interfering RNA may present a tool for hepatic fibrosis treatment.
Oxytocin is released from supraoptic magnocellular neurons (SON) and is thought to act at presynaptic receptors to inhibit transmitter release. Hirasawa, M. et al. showed that WIN55,212-2, a cannabinoid receptor agonist, mimicked the action of oxytocin and occluded oxytocin-induced presynaptic inhibition7. AM251, a CB1 receptor antagonist, blocked both the WIN55,212-2 and the oxytocin-induced presynaptic inhibition of EPSCs. Depolarization of postsynaptic magnocellular neurons caused a transient inhibition of EPSCs that could be blocked by both AM251 and Manning compound, an oxytocin/vasopressin receptor antagonist. Alomone Labs Tetrodotoxin (with citrate) was used throughout the study in order to block out TTX-sensitive NaV channels. The presented results provide evidence that exogenous cannabinoids can act at presynaptic inhibitory receptors on afferent excitatory terminals in the SON. Furthermore, depolarization of magnocellular neurons caused the release of endocannabinoids that acted as retrograde messengers.
CB2 Receptors
Cannabinoid (CB) receptors may be involved in the control of bladder function6. Immunohistochemical staining using Anti-Cannabinoid Receptor 2 (extracellular) Antibody (#ACR-003) detected CB2 receptors in rat urothelium and in sensory and cholinergic bladder nerves. Cannabinor, a selective CB2 receptor agonist, increased urination intervals and increased threshold and flow pressures. The results of the current study show that Cannabinor have significant effects on ‘‘afferent’’ urodynamic parameters, and increase bladder capacity in normal awake rats during cystometry. Considering that CB2 receptors are localized on sensory nerves and on the urothelium, peripheral CB2 receptors may be involved in the regulation of mechanoafferent functions of the rat bladder.
References
- Begg, M. et al. (2001) J. Physiol. 531, 95.
- Chen, S.W. et al. (2012) PLoS One 7, e50850.
- Farquhar-Smith, W.P. et al. (2000) Mol. Cell. Neurosci. 15, 510.
- Freund, T.F. et al. (2003) Physiol. Rev. 83, 1017.
- Gonzalez-Islas, C. et al. (2012) J. Neurosci. 32, 13597.
- Gratzke, C. et al. (2010) Eur. Urol. 57, 1093.
- Hirasawa, M. et al. (2004) J. Physiol. 559, 611.
- Hohmann, A.G. et al. (1999) J. Neurophysiol. 81, 575.
- Jennings, E.A. et al. (2001) J. Physiol. 534, 805.
- Martin, W.J. et al. (1999) Brain Res. 822, 237.
- Matsuda, L.A. et al. (1990) Nature 346, 561.
- McDonald, N.A. et al. (2007) Mol. Pharmacol. 71, 976.
- Ohno-Shosaku, T. et al. (2012) Neuroscientist 18, 119.
- Reichenbach, V. et al. (2012) J. Pharmacol. Exp. Ther. 340, 629.
- Sugiura, T. et al. (1996) Biochem. Biophys. Res. Commun. 229, 58.