The “GABAergic” system is the main inhibitory network in the central nervous system and as such influences almost every aspect of neuronal processing and resulting behavior. γ-amunobutiric acid (GABA), released in a typical vesicular synaptic manner from gabaergic neurons, binds to GABA (A) postsynaptic ligand-gated ion channels to mediate the fast electrical inhibitory action of the transmitter. GABA also binds in and around the synapse to a G-protein coupled receptor (GPCR), GABA (B), the topic of this short review, which mediates other feedback or modulatory, usually slower, cellular and systemic responses. Alomone Labs offers various antibodies directed to both ionotropic and metabotropic GABA receptors enabling the advancement of research in this field.
Introduction
GABA (B) receptors are expressed throughout the nervous system, especially in the brain. They are located both on the postsynaptic membrane alongside the GABA (A) ligand-gated ion channels and in presynaptic membranes, where their activation leads to diminished transmitter release. The latter could be either autoreceptors on the GABA releasing presynaptic terminal or heteroreceptors on neighboring terminals or neurons1. GABA (B) receptors on the presynaptic membrane mostly exert their action by releasing the βγ subunits from their G-protein complex, to directly inhibit the typical presynaptic voltage gated Ca2+ channels (Cav2.1 and Cav2.2 or P/Q type and N-type channels)7,9, which normally conduct the necessary Ca2+ for synaptic vesicle fusion and transmitter release. This mechanism might serve as a negative feedback on GABA release, controlled by the amount already released (detected by autoreceptors) as well as a part in a synergistic action to inhibit the excitatory signal (detected by heteroreceptors)1. The latter could be inhibition of glutamate release from a neighboring neuron which is additive to the postsynaptic GABA (A) activation. Another synergistic action of GABA is manifested by the activation of GABA (B) receptors localized on postsynaptic sites. Binding of GABA to these receptors leads to the activation of G-protein activated Inward Rectifier K+ (GIRK or Kir3) channel6, hyperpolarization and postsynaptic inhibition1.
Molecular Structure of GABA (B) Receptors
The basic functional receptor is a heterodimer composed of two similar subunits each with a seven transmembrane α-helix (7 TM) topology, GABA (B) R1 and GABA (B) R28,12. Both subunits have a long extracellular N-terminal, are similar in size (961 and 941 amino acids in human for GABA (B) R1 and GABA (B) R2, respectively), share 54% similarity in their amino acid sequence and are dimerized by a coiled coil domain in their intracellular C-termini3. However, while GABA (B) R1 binds to agonists and initiates a conformational change in the receptor complex, GABA (B) R2 interacts with and transmits this signal to the intracellular G-protein trimer3,12. In addition to their interaction with G-proteins, which is discussed below, it was recently published that native GABA (B) receptors are in complex with intracellular auxiliary subunits of the KCTD (K+ channel tetramerization domain) protein family. Several members of this protein family were coimmunoprecipitated with GABA (B) R1 and GABA (B) R2 from mouse and rat brain membranes and were shown to influence parameters of GABA (B) signaling (such as agonist potency, receptor desensitization etc.) in an isoform specific manner13. This recent finding might explain the variability in native GABA (B) mediated responses, which could not be attributed so far to molecular isoforms, as not many physiologically relevant splice variants of GABA (B) R1 are found. In humans, the two most studied and abundant splice variants are GABA (B) R1a and GABA (B) R1b, which lacks a segment in the N-terminal, giving rise to a shorter extracellular domain1,3,10,13 (see Figure 1 which summarizes schematically GABA (B) receptors5).
Downstream of the Receptor
The most abundant responses to GABA (B) receptor activation are their coupling to G-protein complexes containing a pertussis toxin sensitive Gi/o family member and the β and γ subunits which then dissociate from the complex to bind proteins in their vicinity and modulate their function, in part leading to the inhibition of adenylate cyclase, thereby decreasing cAMP levels1,3,10. Gβγ directly binds to Cav2.1 and Cav2.2 channels making them much less responsive to electrical stimulation7. This molecular event is at the basis of the presynaptic inhibitory action of GABA and is probably due to the clustering of GABA (B) receptors and Cav channels in the presynaptic terminal membrane. Gβγ also binds to the Kir3 subfamily of K+ channels, probably to directly gate it open, allowing K+ efflux and enabling hyperpolarization of the membrane potential6. The GABA (B)-Kir3 system is localized mostly to postsynaptic membranes and its activation maintains a slow synaptic inhibition. GABA (B) was also found to modulate NMDA channels via the PKA pathway as demonstrated by recording synaptic responses in individual postsynaptic dendritic spines4.

A) Functional GABA (B) receptors are formed by the interaction of GABA (B) R1 and GABA (B) R2 receptors through coiled-coil motifs in the C-terminus, and through transmembrane and extracellular domains. Ligand binding is achieved through GABA (B) R1 while GABA (B) R2 undergoes allosteric modulation and is responsible for activating the G-protein. Two GABA (B) R1 isoforms, GABA (B) R1a and GABA (B) R1b, are expressed in the brain. Two “Sushi motifs” are present GABA (B) R1a and absent in GABA (B) R1b. B) In the hippocampus, GABA (B) receptors are located presynaptically, postsynaptically and on extrasynaptic membranes. On GABA releasing terminals, presynaptic GABA (B) autoreceptors inhibit the release of GABA and heteroreceptors inhibit the release of several other neurotransmitters (glutamate for example). On the postsynaptic membrane, GABA (B) receptors activate K+ channels and induce slow inhibitory postsynaptic potentials that inhibit GABA (A) receptor activity. Receptors on extrasynaptic membranes are probably activated by a GABA overflow from nearby synapses.
Adapted from reference 5 with permission of Elsevier.

Immunohistochemical staining of mouse hippocampus frozen sections using Anti-GABA (B) R1 (extracellular) antibody (#AGB-001), (1:100). A. GABA (B) R1 staining (green) is detected in neurons in the CA3 field and in the dentate granule layer (short arrows), as well as in dendrites of CA3 pyramidal neurons (long arrows). B. Staining with mouse anti-GAP43 antibody (red) sets apart the stratum lacunosum moleculare (SLM). C. Confocal merge suggests the presence of GABA (B) R1 in pyramidal neurons.
Experimental procedure and figure processed at Alomone Labs.
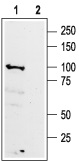
1. Anti-GABA (B) R1 (extracellular) antibody (#AGB-001), (1:200).2. Anti-GABA (B) R1 antibody, preincubated with the negative control antigen.
Experimental procedure and figure processed at Alomone Labs.
Physiology, Pathophysiology and Pharmacology of GABA (B) Receptors
Pharmacologically, GABA (B) receptors are activated by agonists that will eventually lead to specific synaptic inhibition. Therefore, the development of specific agonists may be useful in treating conditions related to hyperexcitability, such as pain. Antagonists on the other hand block endogenous inhibitory actions of GABA and lead to greater activity of specific synapses10. Baclofen, a potent agonist of GABA (B) receptors, was shown to be GABA (B) specific, since its action is absent in the receptor knockout mice strain. As such, it is an important pharmacological tool for differentiating between the metabotropic and ionotropic actions of GABA1,3,10. Baclofen is used in the clinic to treat some forms of spasticity and pain as the compound possesses muscle relaxant properties. This action is carried out probably via inhibitory action on transmitter release in the spinal cord3,10. In addition, subcutaneous administration of baclofen reverses neuropathic hyperalgesia in mice models.
Other agonists of GABA (B) receptors vary in potency, selectivity and the mechanisms they affect. Antagonists are also being developed and are used both as potent research tools and in preclinical trials to treat disorders such as depression or epilepsy3,5,10.
GABA (B) receptors are highly expressed in the limbic system of the brain suggesting their possible involvement in emotional behaviors via presynaptic inhibition2,5. Such implication is suggested for the induction of sound conditioned fear in animals. The complex neuronal circuits leading to such behavior was elegantly demonstrated by the GABAergic inhibitory system which controls in a very defined manner the induction of a specific form of LTP in specific synapses in the amygdala11. The contribution of GABA (B) receptors to neuronal activity was also demonstrated in the hippocampus where the inhibitory activity of GABA (B) receptors is dependent on the stimulation frequency and controls the development of GABAergic synapses in this brain region14.
All these findings nicely demonstrate the important influences on behavior of mammals of a “secondary” GPCR dependent synaptic system, which is used to dynamically and specifically modulate synaptic connectivity and related brain functions.
Alomone Labs offers antibodies to the two GABA (B) Receptors: Anti-GABA (B) R1 (extracellular) (#AGB-001) and Anti-GABA (B) R2 (#AGB-002) antibodies. These antibodies enable the detection of their respective receptors using western blot and immunohistochemistry applications.
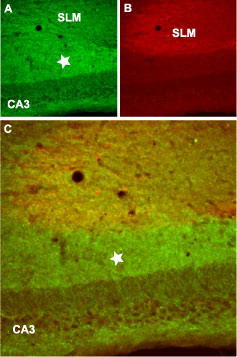
Immunohistochemical staining of mouse hippocampus frozen sections using Anti-GABA (B) R2 antibody (#AGB-002), (1:100). A. GABA (B) R2 staining (green) is detected in layers including the stratum lacunosum moleculare (SLM) and stratum radiatum of the CA3 layer (star). B. Staining with mouse anti-GAP43 antibody (red) sets apart the SLM. C. Confocal merge suggests the presence of GABA (B) R2 in layers, but not in pyramidal neurons.
Experimental procedure and figure processed at Alomone Labs.

1. Anti-GABA (B) R2 antibody (#AGB-002), (1:200).2. Anti-GABA (B) R2 antibody preincubated with the negative control antigen.
Experimental procedure and figure processed at Alomone Labs.
References
- Bettler, B. et al. (2004) Physiol. Rev. 84, 835.
- Bolshakov, V.Y. (2009) Neuron 61, 817.
- Bowery, N.G. et al. (2002) Pharmacol. Rev. 54, 247.
- Chalifoux, J.R. and Carter, A.G. (2010) Neuron 66, 101.
- Cryan, J.F. and Kaupmann, K. (2005) Trends Pharmacol. Sci. 26, 36.
- Dascal, N. (2001) Trends Endocrinol. Metab. 12, 391.
- Dolphin, A.C. (2003) Pharmacol. Rev. 55, 607.
- Jones, K.A. et al. (1998) Nature 396, 674.
- Mintz, I.M. and Bean, B.P. (1993) Neuron 10, 889.
- Ong, J. and Kerr, D.I. (2005) CNS Drug Rev. 11, 317.
- Pan, B.X. et al. (2009) Neuron 61, 917.
- Rondard, P. et al. (2008) EMBO J. 27, 1321.
- Schwenk, J. et al. (2010) Nature 465, 231.
- Xu, C. et al. (2008) Nat. Neurosci. 11, 1410.