Overview
- Peptide (C)SPRVLFSTQPPPTSSDTLDLD, corresponding to amino acid residues 84-104 of rat NGF (precursor) (Accession P25427). Pro-domain of the NGF protein.

 Western blot analysis of Recombinant mouse proNGF protein (#N-250), (lanes 1-3) and Recombinant human proNGF protein (#N-280), (lanes 4-6):1,4. 100 ng + Anti-proNGF Antibody (ANT-005), (1:200).
Western blot analysis of Recombinant mouse proNGF protein (#N-250), (lanes 1-3) and Recombinant human proNGF protein (#N-280), (lanes 4-6):1,4. 100 ng + Anti-proNGF Antibody (ANT-005), (1:200).
2,5. 250 ng + Anti-proNGF Antibody (1:200).
3,6. 500 ng + Anti-proNGF Antibody (1:200).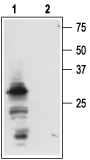 Western blot analysis of mouse submandibular glands (SMGs):1. Anti-proNGF Antibody (#ANT-005), (1:200).
Western blot analysis of mouse submandibular glands (SMGs):1. Anti-proNGF Antibody (#ANT-005), (1:200).
2. Anti-proNGF Antibody, preincubated with proNGF Blocking Peptide (#BLP-NT005).
 Expression of proNGF in rat brain sectionsImmunohistochemical staining of proNGF in rat brain sections using Anti-proNGF Antibody (#ANT-005). A. Staining of astrocytes was performed using mouse anti-glial fibrillary acidic protein (GFAP) (red). B. Co-localization between GFAP and proNGF (orange). C. Staining of proNGF (green).
Expression of proNGF in rat brain sectionsImmunohistochemical staining of proNGF in rat brain sections using Anti-proNGF Antibody (#ANT-005). A. Staining of astrocytes was performed using mouse anti-glial fibrillary acidic protein (GFAP) (red). B. Co-localization between GFAP and proNGF (orange). C. Staining of proNGF (green).
- Lessman, V. et al. (2003) Prog. Neurobiol. 69, 341.
- Bibel, M. and Barde, Y.A. et al. (2000) Genes Dev. 14, 2929.
- Lee, R. et al. (2001) Science 294, 1945.
- Ibanez, C.F. (2002) Trends Neurosci. 25, 284.
- Hasan, W. et al. (2003) J. Neurobiol. 57, 38.
- Beattie, M.S. et al. (2002) Neuron 36, 375.
- Fahnestock, M. et al. (2001) Mol. Cell. Neurosci. 18, 210.
- Lu, B. (2003) Neuron 39, 735.
Neurotrophins are synthesized as pro-forms that can be cleaved either intracellularly to release mature, secreted ligands, or extracellularly by various proteases such as plasmin, furin, PC1/3, PC7, and PACE 4.1,2,5 The immature precursor has a prodomain of 103 amino acids, which was thought to have a role in the folding and sorting of the mature NGF into the various secretion pathways.
It was recently reported that proNGF, binds p75NTR receptor preferentially over TrkA, and this selective binding of proNGF to p75NTR leads to apoptotic death of cells that express both TrkA and p75NTR. However, mature NGF binds and activates both receptors, with resulting promotion of cell survival due to the TrkA-mediated survival signal overriding p75NTR -mediated apoptotic signal.3,4
Since pro- and mature neurotrophins seem to elicit opposite functional effects, by differential interactions with Trks and p75NTR receptors, extracellular cleavage represents a new way to control the synaptic functions of neurotrophins.
It was demonstrated that proNGF from injured spinal cord extracts, is active and induce apoptosis among oligodendrocytes, and apoptosis can be blocked by a proNGF-specific antibody.6
Finally, proNGF was demonstrated as the predominant form in mouse, rat, and human brain tissue, thyroid, hippocampus, thus suggesting a role for proNGF in vivo.5,7,8
Alomone Labs is pleased to offer a highly specific antibody directed against rat proNGF. Anti-proNGF Antibody (#ANT-005) can be used in western blot and immunohistochemical applications. The antibody specifically reacts with proNGF. It has been designed to recognize proNGF from mouse, rat and human samples.
Related products for neutralizing proNGF activity:
Mouse NGF/proNGF Neutralizing Antibody (#ALM-006) can be used in indirect ELISA and neutralization assays to block the biological effects of both NGF and proNGF.
Applications
Citations
 Powered by Bioz
Powered by Bioz- Rat microglia and spinal cord lysates (1:200).
Jung, G.Y. et al. (2013) Glia 61, 1807. - Mouse retina lysate.
Mysona, B.A. et al. (2013) Diabetologia 56, 2329.
- Mouse retina sections (1:100).
Mysona, B.A. et al. (2013) Diabetologia 56, 2329.
- Matragoon, S. et al. (2012) Mol. Vis. 18, 2993.
- Peleshok, J.C. and Ribeiro da Silva, A. (2012) Mol. Pain 8, 1.
- Shulga, A. et al. (2012) J. Neurosci. 32, 1757.
- Al-Gayyar, M.M. et al. (2011) Diabetologia 54, 669.
- Spinnler, K. et al. (2011) J. Biol. Chem. 286, 31707.
- Weinstock, M. et al. (2011) Neurobiol. Aging 32, 1069.
- Ali, T.K. et al. (2011) Diabetologia 54, 657.
- Al-Shawi, R. et al. (2008) Eur. J. Neurosci. 27, 2103.
- Watanabe, T. et al. (2008) J. Neurosci. Res. 86, 3566.
- Yune, T.Y. et al. (2007) J. Neurosci. 27, 7751.
- Bruno, M.A. and Cuello, C.A. (2006) Proc. Natl. Acad. Sci. U.S.A. 103, 6735.
