Overview
- Peptide (C)DYLRPPSTDMNGK, corresponding to amino acid residues 478 - 490 of mouse SORCS2 (Accession Q9EPR5). Extracellular, N-term.

 Western blot analysis of rat brain lysate (lanes 1 and 3) and mouse brain lysates (lanes 2 and 4):1-2. Anti-SORCS2 (extracellular) Antibody (#ANR-135), (1:200).
Western blot analysis of rat brain lysate (lanes 1 and 3) and mouse brain lysates (lanes 2 and 4):1-2. Anti-SORCS2 (extracellular) Antibody (#ANR-135), (1:200).
3-4. Anti-SORCS2 (extracellular) Antibody, preincubated with SORCS2 (extracellular) Blocking Peptide (BLP-NR135).
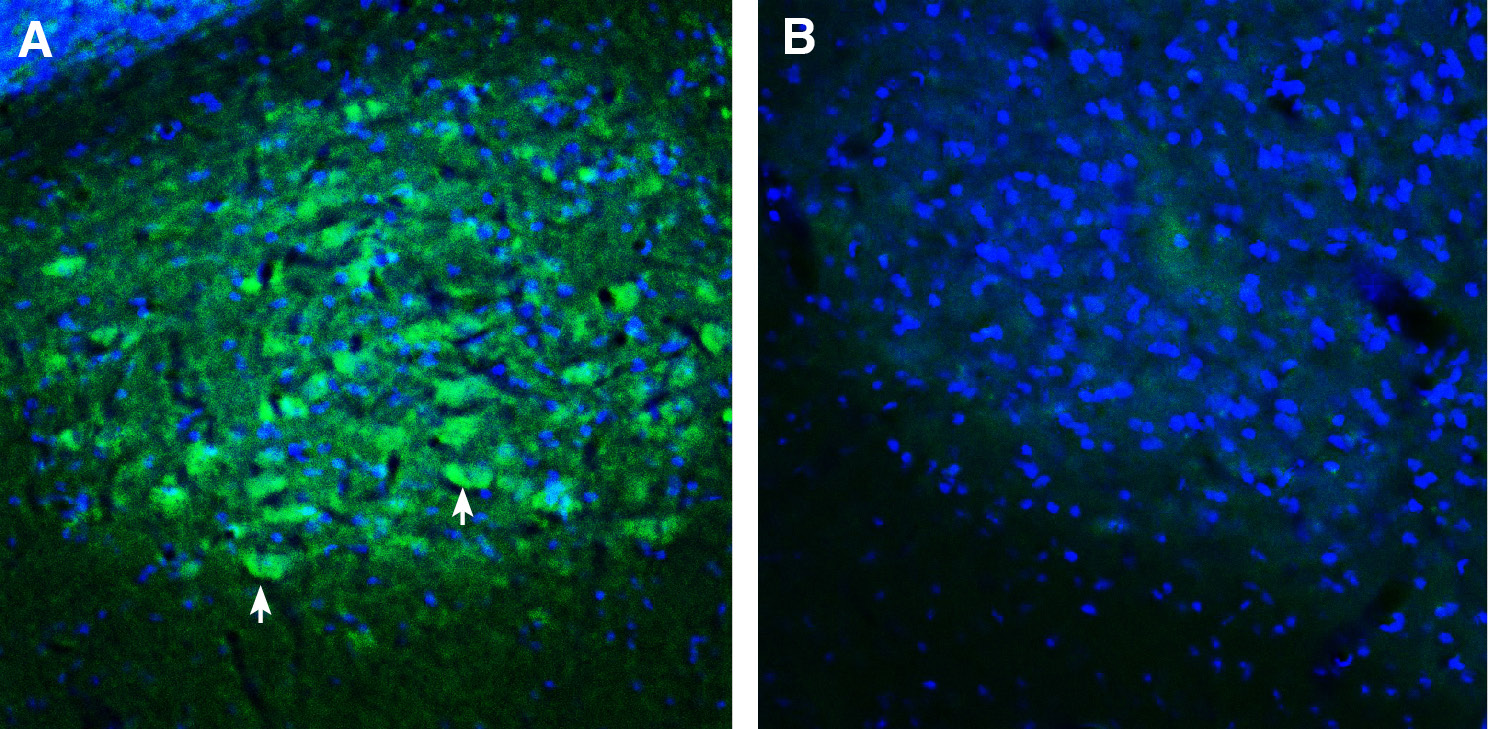 Expression of SORCS2 in mouse deep cerebellar nuclei.Immunohistochemical staining of perfusion-fixed frozen mouse brain sections with Anti-SORCS2 (extracellular) Antibody (#ANR-135), (1:300), followed by goat anti-rabbit-AlexaFluor-488. A. SORCS2 immunoreactivity (green) appears in neuronal outlines (arrows). B. Pre-incubation of the antibody with SORCS2 (extracellular) Blocking Peptide (BLP-NR135), suppressed staining. Cell nuclei are stained with DAPI (blue).
Expression of SORCS2 in mouse deep cerebellar nuclei.Immunohistochemical staining of perfusion-fixed frozen mouse brain sections with Anti-SORCS2 (extracellular) Antibody (#ANR-135), (1:300), followed by goat anti-rabbit-AlexaFluor-488. A. SORCS2 immunoreactivity (green) appears in neuronal outlines (arrows). B. Pre-incubation of the antibody with SORCS2 (extracellular) Blocking Peptide (BLP-NR135), suppressed staining. Cell nuclei are stained with DAPI (blue). Expression of SORCS2 in rat hippocampus.Immunohistochemical staining of perfusion-fixed frozen mouse brain sections with Anti-SORCS2 (extracellular) Antibody (#ANR-135), (1:300), followed by donkey anti-rabbit-biotin and Streptavidin-Cy3. A. SORCS2 immunoreactivity (red) appears in CA1 cells (arrows). B. Pre-incubation of the antibody with SORCS2 (extracellular) Blocking Peptide (BLP-NR135), suppressed staining. Cell nuclei are stained with DAPI (blue).
Expression of SORCS2 in rat hippocampus.Immunohistochemical staining of perfusion-fixed frozen mouse brain sections with Anti-SORCS2 (extracellular) Antibody (#ANR-135), (1:300), followed by donkey anti-rabbit-biotin and Streptavidin-Cy3. A. SORCS2 immunoreactivity (red) appears in CA1 cells (arrows). B. Pre-incubation of the antibody with SORCS2 (extracellular) Blocking Peptide (BLP-NR135), suppressed staining. Cell nuclei are stained with DAPI (blue).
- Januliene, D., et al., (2017) J Mol Biol, 429(19): p. 2907-2917
- Malik, A.R. and T.E. Willnow, (2020) Trends Neurosci, 43(11): p. 870-885.
- Hermey, G., et al., (2004) J Neurochem, 88(6): p. 1470-6.
- Binzer, S., et al.(2016) Mult Scler, 22(6): p. 733-40.
- Christoforou, A., et al., (2011) Mol Psychiatry, 16(3): p. 240-2.
- Ollila, H.M., et al.,(2009) Mol Psychiatry, 14(4): p. 351–353.
- Rezgaoui, M., et al., (2001) Mech Dev, 100(2): p. 335-8.
- Deinhardt, K., et al.,(2011) Sci Signal, 4(202): p. ra82.
- Mori, F., et al.(2015), Neurosci Lett, 608: p. 6-11.
- Baum, A.E., et al.(2008), Mol Psychiatry, 13(2): p. 197-207.
- Anastasia, A., et al.,(2013) Nat Commun, 4: p. 2490.
- Glerup, S., et al., (2016) Mol Psychiatry, 21(12): p. 1740-1751.
- Abdul Aziz, N.A., et al., (2016) BMC Med Genomics, 9(1): p. 58.
- Oelsner, K.T., et al., (2017) BMC Genomics, 18(1): p. 57.
- Wei, W.H., et al., (2012) Eur J Hum Genet, 20(8): p. 857-62.
- Boggild, S., et al., (2016) BMC Cell Biol, 17: p. 8.
- Westergaard, U.B., (2005) FEBS Lett, 579(5): p. 1172-6.
- Glerup, S., et al., (2014) Neuron, 82(5): p. 1074-87.
SORCS2 is a member of the Vps10p-domain receptor family, which comprises five multifunctional neuronal receptors, type I transmembrane receptors Sortilin in humans or sortilin in mice (encoded by the gene SORT1 or Sort1, respectively), SORLA/SorLA (encoded by SORL1/Sorl1) and SORCS1-3/SorCS1-3 (encoded by SORCS1- 3/Sorcs1-3)1, 2. The receptor family is expressed predominantly in the nervous system (both developing and adult) and involved in intracellular sorting and trafficking of various neurotrophic factors and their precursor forms, as well transmembrane receptors and synaptic proteins1.
This gene family has been implicated in a broad range of cellular processes, including neuronal health, differentiation and synaptic plasticity1, 2. The SorCS subfamily contains three closely related members, with at least 49% identity between the VPS10 domains3. SorCS members also contain a leucine rich region (LRR), with 63% identity between SorCS1 and SorCS3 versus only 35% identity between SorCS1 and SorCS23. The three SorCS genes are differentially expressed and regulated in mature neurons3.
Since their discovery a growing number of studies have linked SorCS genes to diverse neurodegenerative diseases such as autism, schizophrenia, bipolar disorder, attention deficit hyperactivity disorder, late-onset Alzheimer disease and multiple sclerosis4-6. SorCS2 is the VPS10 family member that is the least characterized. Ten years after its identification7, a first role for SorCS2 in proneurotrophin-mediated neurodegeneration as co-receptor of the p75NTR was uncovered8. Roles for SorCS2 in amyotrophic lateral sclerosis, bipolar disorder and schizophrenia have been suggested5, 6, 9, but the mechanism of action of SorCS2 is poorly understood10, though recent studies have shown that SorCS2 regulates pro-BDNF dependent synapse plasticity11, 12. Furthermore, SorCS2 has been linked to obesity and colorectal cancer, suggesting a role akin to Sortilin’s for SorCS2 in these diseases13-15.
SorCS2 is localized predominantly in the brain, but also in heart, liver and kidney7, 16. Unlike Sortilin, SorCS2 is preferentially located at the cell surface in mature neurons of the central nervous system (CNS) and glia of the peripheral nervous system (PNS), making it the preferred co-receptor for p75-dependent apoptosis17. SorCS2 contains a leucine-rich region linking the VPS10 domain and the transmembrane domain. It has been shown to exist both as a single chain receptor in the neurons of the CNS and as a cleaved two-chains receptor in the glia of the PNS18. The proteolytic processing site yielding this two-chain version of SorCS2 is located in the leucine-rich region at residue S1031. SorCS2 interacts with proneurotrophins8, 11, 12. Both single and double-chain SorCS2 interact with proneurotrophins (and with less affinity to mature neurotrophins) and form a ternary complex with p75NTR, but the single chain receptor induces growth cone collapse of dopaminergic neurons while the double-chain receptor transduces apoptosis signals in PNS glia18.
Anti-SORCS2 (extracellular) Antibody (#ANR-135) is a highly specific antibody directed against an epitope of the mouse protein. The antibody can be used in western blot and immunohistochemistry applications. It has been designed to recognize SORCS2 from rat, mouse and human samples.
 Powered by Bioz
Powered by Bioz