Add $870 to your cart to get your free sample.
Overview
ω-Conotoxin GVIA (#C-300) is a highly pure, synthetic, and biologically active peptide toxin.
Alternative Name SNX-124
Structure

For research purposes only, not for human use
Applications

Citations (59)
Citations
 Powered by Bioz
Powered by BiozPublished figures using this product
 Knockdown of FMRP enhances synaptic vesicle exocytosis in presynaptic terminals of DRG neurons via CaV2.2 channels.A and B. vGpH response to 40 AP at 10 Hz from presynaptic terminals of DRG neurons transfected with Ctrl shRNA (A) or FMRP shRNA (B) before and after treatment with ω-Conotoxin GVIA (#C-300) and ω-Agatoxin IVA (#STA-500). Fluorescence intensities were normalized to the peak of a brief application of NH4Cl. C. Normalized vGpH responses to 40 AP at 10 Hz from presynaptic terminals of DRG neurons transfected with Ctrl shRNA (black-filled bar, 100±10.6%, n = 38) or FMRP shRNA (red open bar, 137.0±12.6%, n = 25, P = 0.027). ω-Conotoxin GVIA (ConoTx) reduces Ctrl shRNA and FMRP shRNA responses to a similar level (44.7±4.9%, n = 15 and 41.6±3.3%, n = 24, respectively). ω-Conotoxin GVIA and ω-Agatoxin IVA (AgaTx) application further reduces the responses: Ctrl shRNA = 17.3±3.2%, n = 38, and FMRP shRNA = 18.1±5.0%, n = 27. D. Average vGpH response to a 40 Hz stimulation for 30 s from presynaptic terminals of DRG neurons transfected with Ctrl shRNA (blackfilled squares) or FMRP shRNA (open red squares).
Knockdown of FMRP enhances synaptic vesicle exocytosis in presynaptic terminals of DRG neurons via CaV2.2 channels.A and B. vGpH response to 40 AP at 10 Hz from presynaptic terminals of DRG neurons transfected with Ctrl shRNA (A) or FMRP shRNA (B) before and after treatment with ω-Conotoxin GVIA (#C-300) and ω-Agatoxin IVA (#STA-500). Fluorescence intensities were normalized to the peak of a brief application of NH4Cl. C. Normalized vGpH responses to 40 AP at 10 Hz from presynaptic terminals of DRG neurons transfected with Ctrl shRNA (black-filled bar, 100±10.6%, n = 38) or FMRP shRNA (red open bar, 137.0±12.6%, n = 25, P = 0.027). ω-Conotoxin GVIA (ConoTx) reduces Ctrl shRNA and FMRP shRNA responses to a similar level (44.7±4.9%, n = 15 and 41.6±3.3%, n = 24, respectively). ω-Conotoxin GVIA and ω-Agatoxin IVA (AgaTx) application further reduces the responses: Ctrl shRNA = 17.3±3.2%, n = 38, and FMRP shRNA = 18.1±5.0%, n = 27. D. Average vGpH response to a 40 Hz stimulation for 30 s from presynaptic terminals of DRG neurons transfected with Ctrl shRNA (blackfilled squares) or FMRP shRNA (open red squares).
Adapted from Ferron, L. et al. (2014) with permission of Springer Nature.
Electrophysiology
- Rat INS-1 832/12 cells (whole-cell calcium currents).
Luan, C. et al. (2019) Commun. Biol. 2, 106. - Rat DRGs (patch clamp).
Moutal, A. et al. (2018) Neuroscience 381, 79.
Product citations
- Torturo, C.L. et al. (2019) eNeuro 6, e0278.
- Lubbert, M. et al. (2017) eLife 6, e28412.
- Forostyak, O. et al. (2016) Stem Cell Res. 16, 622.
- Grimsley, C.A. et al. (2016) J. Neurophysiol. 116, 2550.
- Luster, B.R. et al. (2016) Physiol. Rep. 4, e12787.
- D’Onofrio, S. et al. (2016) Physiol. Rep. 4, e12740.
- Pennock, R.L. and Hentges, S.T. (2016) J. Neurophysiol. 115, 2376.
- Squecco, R. et al. (2016) Mol. Cell. Neurosci. 75, 50.
- D’Arco, M. et al. (2015) J. Neurosci. 35, 5891.
- Gan, K.J. et al. (2015) Mol. Biol. Cell 26, 1058.
- Lopez Soto, E.J. et al. (2015) J. Gen. Physiol. 146, 205.
- Cassidy, J.S. et al. (2014) Proc. Natl. Acad. Sci. U.S.A. 111, 8979.
- Ferron, L. et al. (2014) Nat. Commun. 5, 3628.
- Gomez-Sanchez, R. et al. (2014) Neurobiol. Dis. 62, 426.
- Hernandez-Gonzalez, O. et al. (2014) Purinergic Signal. 10, 269.
- Perez-Burgos, A. et al. (2014) FASEB J. 28, 3064.
- Tu, H. et al. (2014) Am. J. Physiol. 306, C132.
- Alamilla, J. and Gillespie, D.C. (2013) PLoS ONE 8, e75688.
- Amato, A. et al. (2013) Eur. J. Pharmacol. 718, 131.
- Eggers, E.D. et al. (2013) J. Neurophysiol. 110, 709.
- Izquierdo Serra, M. et al. (2013) Biochim. Biophys. Acta. 1830, 2853.
- Jones, S.L. and Stuart, G.J. (2013) J. Neurosci. 33, 19396.
- Kim, S.H. and Ryan, T.A. et al. (2013) J. Neurosci. 33, 8937.
- Lowe, M. et al. (2013) Exp. Physiol. 98, 1199.
- Radzicki, D. et al. (2013) J. Neurosci. 33, 9920.
- Shutov, L.P. et al. (2013) J. Physiol. 591, 2443.
- Sia, T.C. et al. (2013) Am. J. Physiol. 305, G933.
- Tzour, A. et al. (2013) J. Neuroendocrinol. 25, 76.
- Wedemeyer, C. et al. (2013) J. Neurosci. 33, 15477.
- Wen, H. et al. (2013) eLife 2, e01206.
- Wen, H. et al. (2013) J. Neurosci. 33, 7384.
- Abitbol, K. et al. (2012) J. Physiol. 590.13, 2977.
- Baillie, L.D. et al. (2012) Neuropharmacology 63, 362.
- Wang, S. et al. (2012) Cereb. Cortex 2, 584.
- Xiong, Q.J. et al. (2012) Am. J. Physiol. 303, C376.
- Alle, H. et al. (2011) J. Neurosci. 31, 8001.
- Martel, P. et al. (2011) PLoS ONE 6, e20402.
- Craviso, G.L. et al. (2010) Cell. Mol. Neurobiol. 30, 1259.
- Meunier, F.A. et al. (2010) J. Cell Sci. 123, 1131.
- Braun, M. et al. (2008) Diabetes 57, 1618.
Specifications
Properties
Technical Specifications
Origin Synthetic peptide
MW 3037 Da
Sequence CKSXGSSCSXTSYNCCRSCNXYTKRCY -NH2
Modifications Disulfide bonds between Cys1-Cys16, Cys8-Cys19 and Cys15-Cys26. X = Hydroxyproline. Tyr27 - C-terminal amidation.
Peptide Content 100%
Purity >99% (HPLC)
Molecular formula C120H188N38O43S6.
CAS No. 106375-28-4
Biological Activity
Target N-type Ca2+ channels
Effective concentration 20 nM - 1 μM.
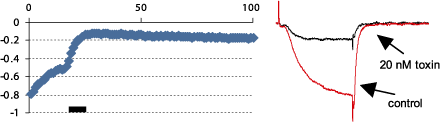
Activity ω-Conotoxin GVIA specifically and reversibly blocks N-type CaV channels1 and is reported to antagonize P2X receptors2.
References
- McCleskey, E.W. et al. (1987) Proc. Natl. Acad. Sci. U.S.A. 84, 4327.
- Lalo, U.V. et al. (2001) Brain Res. Bull. 54, 507.
Solubility and Storage
Shipping and storage Shipped at room temperature. Product as supplied can be stored intact at room temperature for several weeks. For longer periods, it should be stored at -20°C.
Solubility Any aqueous buffer. Centrifuge all product preparations before use (10000 x g 5 min).
Storage of solutions Up to two weeks at 4°C or three months at -20°C.
Scientific Background
References
Scientific background
ω-Conotoxin GVIA is a synthetic toxin originally isolated from the Conus geographus. ω-Conotoxin GVIA is a specific blocker of CaV2.2 Ca2+ channels. It specifically blocks N-type CaV channels by binding to the CaV2.2 α1 subunit (α1B) and its action is only partially reversible.2,3 In accordance, it inhibits synaptic transmission in many systems.4 It is also reported to antagonize P2X receptors.5 The toxin is used to specifically investigate CaV2.2 channel's contributions (by subtraction of the activity before and during perfusion of the toxin). Alternatively, it is used to eliminate the N-type channel contribution to highlight some other channel or enzyme activity. ω-Conotoxin GVIA was also used to purify the channel protein using immunoprecipitation techniques and to label and localize channels and synapses.6
References
- Olivera, B.M. et al. (1984) Biochemistry 23, 5087.
- McCleskey, E.W. et al. (1987) Proc. Natl. Acad. Sci. U.S.A. 84, 4327.
- Feng, Z.P. et al. (2001) J. Biol. Chem. 276, 15728.
- Kerr, L.M. and Yoshikami, D. (1984) Nature 308, 282.
- Lalo, U.V. et al. (2001) Brain Res. Bull. 54, 507.
- Pravettoni, E. et al. (2000) Dev. Biol. 227, 581.
Last Update: 11/03/2025
