
What if we told you that small extracellular vesicles (sEVs) – those little sacks carrying goods around and between cells – coupled with a voltage-dependent potassium (Kv) channel and a heat shock protein, might just help to diagnose cardiovascular diseases (CVDs) like heart failure (HF)? Well, it looks like that might very well be case. In a recent, small-scale study, a team from Chile has put forward the idea of using circulating sEVs and the amount of hERG1 and Hsp47 in their surface as a reliable minimal-invasive tool to diagnose heart conditions (1). It’d be a significant step forward if it turns out to be true, but how did they come to this conclusion and what makes the proteins hERG1 and Hsp47 so significant?
A Closer Look at hERG1 and Hsp47 in Heart Disease
Small extracellular vesicles, which are membrane-bound vesicles that carry proteins, lipids, or nucleic acids between cells, have been gaining more and more attention thanks for their crucial role in intercellular communication. Most relevant here is the research that spotted circulating sEVs in multiple different cardiovascular conditions (2), which of course makes any good scientist think, “Biomarker!”.
Now let’s turn our attention to two proteins of interest, heat shock protein 47 (Hsp47) and the voltage-dependent potassium channel subunit KV11.1 (aka the human ether-a-go-go-related gene 1, or hERG1). We already have in vivo work that nicely demonstrates how sEVs containing Hsp70 offer a degree of cardioprotection by binding toll-like receptor 4 (TLR4) and switching on protective pathways through a different heat shock protein, Hsp27 (3, 4). So, we can see that heat shock proteins already have history in the cardiovascular world. Hsp47 in particular is upregulated in response to cardiac stress and injury – like myocardial infarction or HF – and is involved in cardiac injury-induced fibrosis (5, 6), making it perfect to examine as a possible biomarker of CVD.
Our potassium channel here, hERG1, needs no introduction to anyone working in the cardiac subjects, but just to recap, hERG1 is absolutely essential to normal electrical activity in the heart, and disruptions in hERG1 function – often due to genetic mutations – can lead to long QT syndrome (LQTS), which predisposes individuals to arrhythmias (7, 8). The basic and critical function of hERG1 has put it centre stage in a huge amount of cardiovascular and cardiotoxicity research to date.
Based on this background, the researchers started with a straightforward question: could they detect hERG1 and Hsp47 present on membranes of sEVs from blood samples of patients with heart disease?
To answer this, they isolated sEVs from the blood of patients with ischemic heart. Western blot and flow cytometry results both made use of specific antibodies against hERG1 and Hsp47, including Alomone’s Anti-KCNH2 (HERG) (extracellular) Antibody (#APC-109), to confirm the expression of hERG1 and Hsp47 on the sEVs (Figure 1). The data also show how levels of these proteins were elevated in ischemic patients compared to controls, making it more likely the presence of hERG1 and Hsp47 in sEVs (which we’ll call EV-hERG1 and EV-HSP47) was directly tied to acute cardiac events.


Figure 1. hERG1 (EV-hERG1) and Hsp47 (EV-Hsp47) epitopes are expressed on the surfaces of sEVs from cardiac ischemia patients. (A). Pooled sEV samples from individuals with cardiac ischemia were purified from platelet-free plasma by several ultracentrifugation steps and tested via western blotting using Anti-KCNH2 (HERG) (extracellular) Antibody (#APC-109), anti-Hsp47, and anti-CD9 antibodies. Both the sEV-free fraction (−, “12,000× g pellet”) and the sEV-enriched fraction (+, “110,000× g pellet”) were analyzed. (C). Flow cytometry: purified sEVs from blood plasma enriched by ultracentrifugation were incubated with or without anti-hERG1, then incubated with α-rabbit IgG Alexa Fluor 647 antibodies. (D). Flow cytometry: purified sEVs from blood plasma enriched by ultracentrifugation were incubated with or without anti-Hsp47, then incubated with α-rabbit IgG Alexa Fluor 647 antibodies. Adapted from Osorio et al (2024).
The Role of Hypoxia: What Happens When Cells Are Starved of Oxygen?
The next logical question was, “What triggers the release of these proteins into sEVs?” The researchers hypothesized that hypoxia – an oxygen-deprived state common in heart attacks – might be to blame. To test this theory, they turned to cardiomyocytes and subjected them to a hypoxia stress test in the lab. The results were striking: within just 15 minutes at 1% oxygen, the cells ramped up the release of sEVs, peaking at 30 minutes (Figure 2).
Using our Anti-KCNH2 (HERG) (extracellular) Antibody (#APC-109) in an ELISA, they saw how these sEVs were loaded with hERG1 and Hsp47 – far more than in cells under normal oxygen levels. Together, thess data suggest hypoxic conditions actively drive the heart to release these protein-laden vesicles, potentially as a distress signal. This could mean a simple blood test to detect these proteins might be indicative of cardiac distress, which would allow for early intervention before the damage becomes irreversible

Figure 2. Extracellular vesicles containing hERG1 (EV-hERG1) and Hsp47 (EV-Hsp47) are secreted by cardiomyocytes at low concentrations of oxygen. (A) The sEV concentration was studied in primary cell culture from rat neonatal cardiomyocytes exposed to normoxia (21% oxygen) and hypoxia (1% oxygen) via nanoparticle tracking analysis (NTA). The bar graph represents the mean ± SD (n = 7 per group), and the statistical analysis was conducted using an ANOVA, Tukey’s post hoc test, and the Mann–Whitney Test. Significance is denoted as * p < 0.05 and ** p < 0.005 vs. normoxia, and # p < 0.05 vs. 15 min of exposure to hypoxia. (B) The EV-hERG1 concentration was studied in a human cardiomyocyte cell line (AC16) exposed to normoxia (21% oxygen) and hypoxia (1% oxygen) in an FBS-free culture medium for 1 h using in-house ELISA. The bar graph represents the mean ± SD (n = 6 per group), and the statistical analysis was conducted using the t-test and Mann–Whitney Test. Significance is denoted as * p < 0.05 vs. normoxia. (C) The EV-Hsp47 concentration was studied in a human cardiomyocyte cell line (AC16) exposed to normoxia (21% oxygen) and hypoxia (1% oxygen) in an FBS-free culture medium for 1 h using in-house ELISA. The bar graph represents the mean ± SD (n = 4 per group), and the statistical analysis was conducted using a t-test and Mann–Whitney test. Significance is denoted as * p < 0.05 vs. normoxia. Figure and legend from Osorio et al (2024).
Size Matters: The Diagnostic Power of sEVs
At this point, you’re probably wondering “Is the presence of these proteins enough to make a diagnosis?” That’s a fair question, especially considering the small size of this study. To help answer this, the Chilean team took a closer look at some of the less obvious questions around sEVs as diagnostics. This is why they decided to look at the size of the vesicles that hERG1 and Hsp47 inhabit.
What they noticed was the potential predictive power of EV-hERG1 versus EV-Hsp47 was not uniform across all sEVs. In fact, smaller vesicles, between 60 and 120 nm were particularly rich in hERG1, while those between 120 and 300 nm were more likely to carry Hsp47.
So, why does this matter? This size-specific distribution could help fine-tune a diagnostic tool, making it more accurate or even more discerning. Developing a reliable diagnostic tool wouldn’t just be about detecting the proteins, it would need to encompass the full picture of where and how they appear.

Figure 3. The levels of extracellular vesicles containing hERG1 (EV-hERG1) and Hsp47 (EV-Hsp47) increased in the blood samples during cardiac ischemia. The concentrations of sEVs containing hERG1 or Hsp47 were studied in platelet-free plasma (PFP) from blood samples of the stress test participants using nanoparticle tracking analysis. (A) The sEV concentration was studied in platelet-free plasma from the stress test participants using nanoparticle tracking analysis (NTA). The boxes represent the interquartile range of the values, whereas the whiskers’ spans represent the minima to maxima, showing all points for the control (n = 13), cardiac-ischemia-negative (n = 13), and cardiac-ischemia-positive (n = 13) samples. The red arrow points it out a participant with a negative cardiac ischemia diagnosis but with elevated levels of sEVs in their blood during the stress test. In B and C, the amounts of 1 × 109 sEVs were determined for each blood plasma (PFP) sample and immunolabeled for 1 h with primary antibody, and then for 1 h with Alexa Fluor 532 secondary antibody. The graphic represents the averages for the patients with negative (n = 13) and positive (n = 13) cardiac ischemia diagnoses, as described in Table 1. The statistical analysis was conducted using ANOVA, Tukey’s post hoc test, and the Mann–Whitney Test. Significance is denoted as *** p < 0.0005. (B) anti-hERG1 antibody; (C) anti-Hsp47 antibody. Figure and legend from Osorio et al (2024).
Chronic vs. Acute: How Do These Markers Behave Over Time?
With these findings in hand, the team turned to the bigger question of how hERG1 and Hsp47 levels change in chronic versus acute heart conditions. Here they looked at patients diagnosed with compensated heart failure (CHF) and decompensated heart failure (DHF). What they found added another layer of complexity to the story.
With help from nanoparticle tracking analysis (NTA) and our Anti-KCNH2 (HERG) (extracellular) Antibody (#APC-109), the team built a more nuanced view of how hERG1 and Hsp47 levels in sEVs vary depending on the clinical condition. In all three groups – healthy, CHF, and DHF – there were detectable levels of EV-hERG1 and EV-Hsp47 in their platelet-free plasma. The size distribution of the sEVs was similar across the groups, so we can assume size alone doesn’t differentiate the clinical conditions.
However, the overall concentration of sEVs was higher in the CHF group compared to healthy individuals, while in the DHF group the concentration of sEVs was significantly lower than in both the CHF group and healthy individuals. Looking at our marker proteins specifically reveals a similar picture. Levels of EV-hERG1 were significantly increased in CHF patients compared to healthy individuals, yet in DHF patients, EV-hERG1 levels were reduced by approximately 10-fold compared to both the CHF group and the healthy group. Similar to hERG1, the concentration of EV-Hsp47 was lower in DHF patients than in CHF patients and healthy individuals.
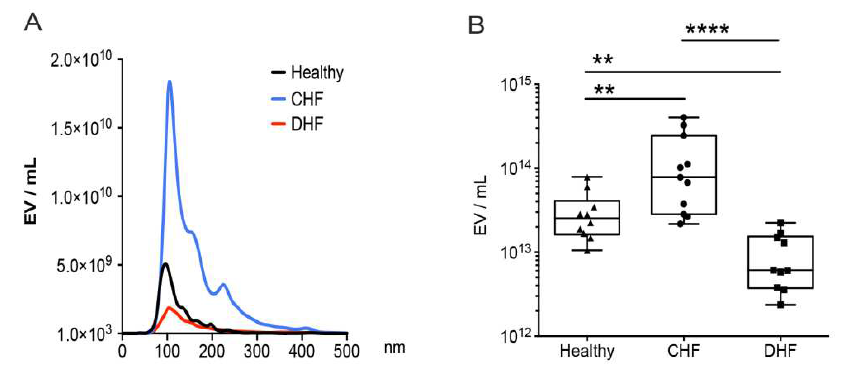

Figure 4. The presence of sEVs with hERG1 and Hsp47 decreased in the blood plasma of patients with decompensated heart failure. Platelet-free plasma from the blood of healthy (n = 10), compensated heart failure (CHF, n = 10), and decompensated heart failure (DHF, n = 10) participants was used to evaluate the concentrations of total sEVs, EV-hERG1, and EV-Hsp47 using the NTA method. (A) sEVs size distributions; (B) plasma sEV concentrations (EV/mL); (C) plasma EV-hERG1 concentrations (EV-hERG1/mL); (D) plasma EV-Hsp47 concentrations (EV-Hsp47/mL). The boxes represent the interquartile ranges of the values, whereas the whiskers’ spans represent the minima to maxima, showing all points for CHF (n = 10) and DHF (n = 10) participants. The statistical analysis was conducted using ANOVA, Tukey’s post hoc test, and the Mann–Whitney test. Significance is denoted as * p < 0.05; ** p < 0.005; *** p < 0.005, and **** p < 0.00005. ns, not significant. Figure and legend from Osorio et al (2024).
What does this all mean? Well, while the levels of EV-hERG1 and EV-Hsp47 increase during acute cardiac events, such as those induced during a stress test (e.g. transient ischemia), they decrease significantly in chronic conditions like DHF. This decrease in sEV, EV-hERG1, and EV-Hsp47 levels in DHF patients could point to a diminished ability of the heart to respond to a chronic stress or a shift in the pathophysiology of heart failure as it progresses from a compensated to a decompensated state.
From a diagnostic perspective, hERG1 and Hsp47 seem to be a potential fit for spotting acute cardiac events, the diagnostic criteria for spotting chronic conditions might need more work. It also highlights the need for different biomarkers to track disease progression over time.
A Glimpse into the Future of Cardiac Diagnostics
So, where does this leave us? The research from Osorio et al. (2024) has built a solid foundation for using EV-hERG1 and EV-Hsp47 as biomarkers for CVDs like HF, being the first group to show that hERG1 and the Hsp47 are present in the membranes of sEVs from human peripheral blood. But like all good science, it also raises new questions: wow exactly do these proteins get into sEVs? Could other markers in sEVs provide even more predictive power? And most importantly, how can these findings be translated into real-world diagnostics?
Studies like this one highlight the need to understand the subtleties of EVs – a topic that perhaps missed out on the focus it deserved. Thankfully, more and more are working to unravel the mechanisms at play here, and at Alomone Labs we plan to keep pressing on with our in-house development of new and reliable antibodies to help with your EVs research. We are specifically proud to present our new series of conjugated antibodies for the EVs markers CD9, CD63, CD81 and calnexin (see below).
It’s early days for hERG1 and Hsp47, but the road ahead is full of potential, and with every assay, every data point, every new idea, we’re one step closer to transforming how we diagnose and treat heart diseases.
Alomone’s fluorescently labeled primary antibodies for EVs markers
References
1. L. A. Osorio, M. Lozano, P. Soto, V. Moreno-Hidalgo, A. Arévalo-Gil, A. Ramírez-Balaguera, D. Hevia, J. Cifuentes, Y. Hidalgo, F. Alcayaga-Miranda, C. Pasten, D. Morales, D. Varela, C. Urquidi, A. Iturriaga, A. Rivera-Palma, R. Larrea-Gómez, C. E. Irarrázabal, Levels of Small Extracellular Vesicles Containing hERG-1 and Hsp47 as Potential Biomarkers for Cardiovascular Diseases. Int J Mol Sci 25, 4913 (2024). DOI: https://doi.org/10.3390/ijms25094913.
2. N. Buntsma, E. van der Pol, R. Nieuwland, A. Gąsecka, Extracellular Vesicles in Coronary Artery Disease. Adv Exp Med Biol 1418, 81–103 (2023). DOI: https://doi.org/10.1007/978-981-99-1443-2_6.
3. S. Saheera, V. P. Jani, K. W. Witwer, S. Kutty, Extracellular vesicle interplay in cardiovascular pathophysiology. Am J Physiol Heart Circ Physiol 320, H1749–H1761 (2021). DOI: https://doi.org/10.1152/ajpheart.00925.2020.
4. A. P. Sklifasovskaya, M. Blagonravov, A. Ryabinina, V. Goryachev, S. Syatkin, S. Chibisov, K. Akhmetova, D. Prokofiev, E. Agostinelli, The role of heat shock proteins in the pathogenesis of heart failure (Review). Int J Mol Med 52, 106 (2023). DOI: https://doi.org/10.3892/ijmm.2023.5309.
5. A. Abdelnasir, J. R. Sun, Y. F. Cheng, H. B. Chen, S. Tang, N. Kemper, J. Hartung, E. D. Bao, Evaluation of Hsp47 expression in heat-stressed rat myocardial cells in vitro and in vivo. Genet Mol Res 13, 10787–10802 (2014). DOI: https://doi.org/10.4238/2014.December.18.20.
6. H. Khalil, O. Kanisicak, R. J. Vagnozzi, A. K. Johansen, B. D. Maliken, V. Prasad, J. G. Boyer, M. J. Brody, T. Schips, K. K. Kilian, R. N. Correll, K. Kawasaki, K. Nagata, J. D. Molkentin, Cell-specific ablation of Hsp47 defines the collagen-producing cells in the injured heart. JCI Insight 4, e128722 (2019). DOI: https://doi.org/10.1172/jci.insight.128722.
7. M. C. Sanguinetti, M. Tristani-Firouzi, hERG potassium channels and cardiac arrhythmia. Nature 440, 463–469 (2006). DOI: https://doi.org/10.1038/nature04710.
8. G. N. Tseng, I(Kr): the hERG channel. J Mol Cell Cardiol 33, 835–849 (2001). DOI: https://doi.org/10.1006/jmcc.2000.1317.